




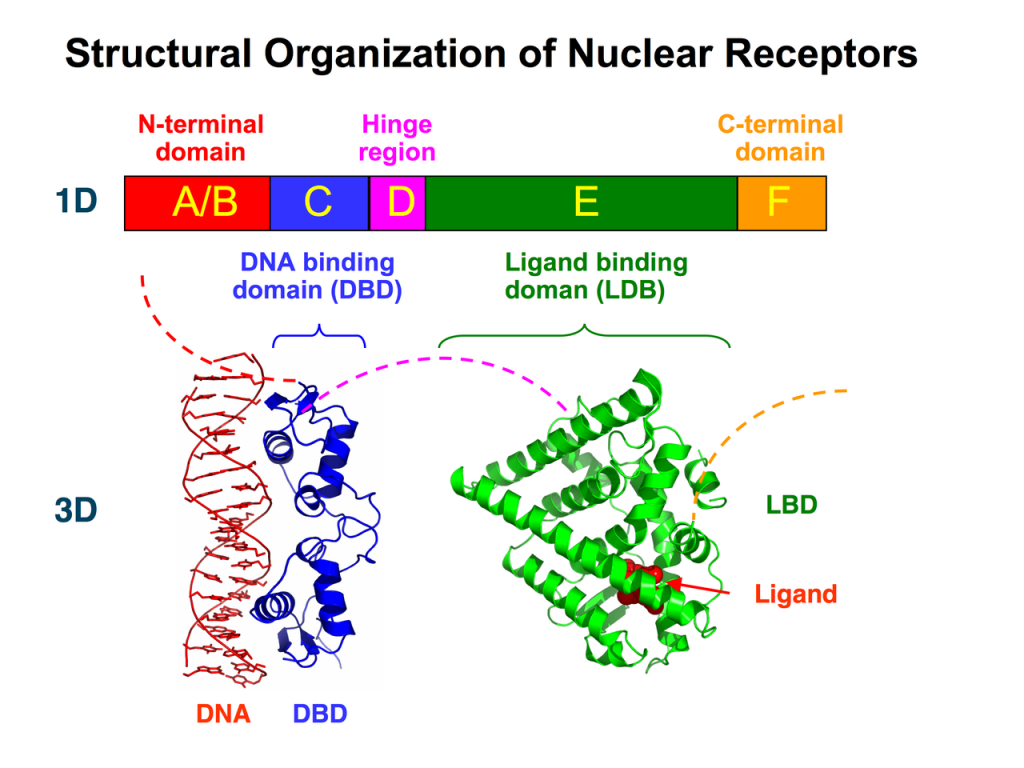
El dominio A/B se sitúa en el extremo N-terminal y es un dominio muy variable; es decir, los dominios A/B de los distintos receptores nucleares pueden ser muy distintos entre sí. Esta variabilidad ya se ve reflejada en su longitud. El dominio A/B puede contener desde 50 a 500 aminoácidos. Sea como sea, curiosamente, suele estar codificado por un solo exón. En este dominio encontramos, como mínimo, una región de transactivación, la AF-1, la cual interactúa con proteínas que determinarán la acción del receptor.
El dominio C es el dominio más conservado entre todos los receptores que lo presentan (de los 48 receptores nucleares, sólo DAX-1 y SHP carecen de él). Esta conservación implica que el dominio C, codificado en dos exones, presenta pocas diferencias entre distintos receptores nucleares. El dominio C codifica para los aminoácidos que conforman el DBD, es decir, para el módulo de reconocimiento del DNA. El DBD presenta un núcleo de 66 aminoácidos con un plegamiento característico (también altamente conservado entre receptores). Este núcleo es responsable tanto del reconocimiento de una secuencia concreta de DNA (se une sólo si encuentra un “sintagma” del DNA concreto) como de la dimerización de los receptores nucleares (muchas veces actúan en parejas). Los “sintagmas” del DNA reconocidas por los receptores nucleares reciben el nombre genérico de Hormone Response Elements (HRE) y suelen estar formados por dos “palabras” separadas por un número determinado de bases (las letras del DNA). Dentro del núcleo de 66 aminoácidos se pueden distinguir secuencias aún más cortas con funciones concretas. Así, la denominada caja P es responsable directa del reconocimiento de la palabra del HRE; la caja D, por su parte, es responsable del reconocimiento de la distancia entre las palabras del HRE.
El dominio D, poco conservado, codifica para una porción flexible de la proteína que sirve de enlace, obviamente, entre la región C y la E; entre el módulo de reconocimiento del DNA (DBD) y el de unión al ligando (LBD). Esta flexión permite a los receptores nucleares adoptar diferentes conformaciones, lo que resulta capital para que la mayoría de ellos actúen por parejas. Además, contiene una señal de localización nuclear, una especie de código postal que indica que esta proteína debe transportarse (según las condiciones) al núcleo.
El dominio E es la región más extensa. Está codificada por 5 exones y se encuentra bastante conservada. Curiosamente, su estructura secundaria (el plegamiento de los aminoácidos que la forman) en 11 o 12 hélices alfa está más conservada que la estructura primaria. Es decir, aunque puedan haber variaciones en los aminoácidos concretos que forman su secuencia lineal, estos se pliegan formando un conjunto de hélices alpha. El dominio E contiene el módulo de unión a ligando (el LBD). Es, por tanto, responsable de la especificidad fisiológica de la acción del receptor: determina en respuesta a qué estímulo actuará. Además, participa en la dimerización de los receptores, en la unión de otras proteínas que modulan su actividad (por ejemplo, coactivadores o heat-shock proteins, si vamos para nota), en su localización en el núcleo y, directamente en su activación dependiente de ligando, ya que presenta el segundo dominio de activación (AF-2).
Este dominio no está presente en todos los receptores y, cuando lo está, presenta una variabilidad muy alta entre ellos.