
| Evolucionismo y psicopatología |
|
|
Recogiendo la tradición platónica, y con una gran influencia posterior sobre la conceptualización de los trastornos mentales, que se mantiene hasta la actualidad, Sir J. Hughlings Jackson (1835-1911) integró la idea de la localización cerebral de las funciones psíquicas con una interpretación incorrecta de la teoría de la evolución basada en la noción de jerarquía.
Los modelos estructurales jerárquicos se aplican en psicología a multitud de constructos. Se han aplicado a la descripción de la inteligencia, también se han utilizado en los estudios sobre temperamento, personalidad y psicopatología. Se encuentran también modelos de estructuras jerárquicas en todas las ciencias, sean éstas físicas, biológicas o sociales. Probablemente ello se deba a que los modelos jerárquicos estrictos, es decir, aquellos que establecen relaciones de control unidireccional desde los niveles superiores, se ajustan bastante bien al concepto clásico de causalidad. En ese sentido, el establecimiento de estructuras jerárquicas mediante el análisis factorial se parece bastante a la experimentación de laboratorio, dado que ésta puede ser vista como una técnica para crear artificalmente un forma simple de jerarquía entre variables. El tema puede complicarse si se entra en la discusión acerca de lo apropiado que pueda ser considerar la relación de control entre niveles como unidireccional o recíproca.
El significado de los modelos estructurales jerárquicos es doble. Por un lado hacen referencia a la organización de un sistema en un momento dado, incluyendo las relaciones de multideterminación recíproca entre sus niveles; éste viene a ser su significado espacial. Así, por ejemplo, los cuerpos celestes se organizan en sistemas, éstos en galaxias, etc.; y las células se organizan en tejidos, éstos en órganos y éstos en organismos, etc. Por otro lado, el significado temporal de los modelos estructurales jerárquicos se refiere a la evolución de la estructura, es decir, a la sucesión de las diferentes etapas de adquisición de niveles. En diferentes campos científicos, las estructuras jerárquicas respectivas tienen sorprendentes similitudes, puesto que se parte de un estado inicial de homogeneidad que, progresivamente, irá sufriendo un proceso de diferenciación. Así, en cosmología se explica la evolución del Universo como un proceso de diferenciación de cuerpos celestes a partir de una singularidad homogénea: el gran estampido que los anglófonos denominan "big-bang". De manera similar, en biología se explica el desarrollo de un organismo como un proceso de división y diferenciación celular a partir de una célula inicial, y los modelos jerárquicos de la inteligencia postulan una inteligencia inicial programada genéticamente para diferenciarse progresivamente en aptitudes específicas a lo largo del desarrollo, con lo que la contribución cualitativa de la herencia y el ambiente queda delimitada.
La idea de jerarquía se ha aplicado también al proceso evolutivo de las especies Según tal noción, éste proceso ha seguido un camino de progresivo perfeccionamiento hasta llegar al Hombre (que se encontraría en la cúspide de la jerarquía evolutiva). Pero esta manera de ver las cosas no se corresponde con la forma en que Darwin entendía la evolución. Para él la evolución era esencialmente oportunista, dando lugar a diferentes formas de adaptación adecuadas al entorno presente en cada momento histórico, pero no ordenadas en virtud de ninguna clase de oscuro principio de perfección o progreso.
 Información complementaria: Neandertales y homo sapiens
Información complementaria: Neandertales y homo sapiens
En cambio, para Spencer la evolución sí que respondía a ese principio teleológico metafísico (Mayr, 1991; Ruse, 1986). Spencer (1862) adoptó la ley de la organización ascendente del Cosmos de von Baer, desde un estado de homogeneidad inicial hacia una progresiva heterogeneidad. Como la evolución del Universo, el desarrollo ontogenético progresa desde la homogeneidad hasta la heterogeneidad, así que la evolución filogenética debería poder caracterizarse también como un proceso en virtud del cual la materia pasa de un estado de homogeneidad indefinida hasta un estado de heterogeneidad definida y coherente.

En el campo de la psicopatología, la concepción de la evolución como progreso reafirmó la idea de que la enfermedad mental era el resultado de una degeneración. De hecho, los estudios epidemiológicos revelaban la existencia de coincidencia en los mismos grupos de individuos de diferentes signos de tal "degeneración", como la enfermedad o el retraso mental, la tuberculosis, el alcoholismo, la criminalidad, altas tasas de mortalidad infantil, pobreza y corta expectativa de vida, indicando que la naturaleza estaba actuando eliminando los individuos genéticamente inferiores. Se argumentaba también que el correlato en la personalidad de esta clase de inferioridad era un síndrome de deficiencia general del carácter (W.B. Maher y B.A. Maher, 1994), con rasgos como la deshonestidad, incapacidad para esperar la gratificación, promiscuidad sexual, holgazanería y otros por el estilo. En las personas con este tipo de personalidad cabía esperar con cierta seguridad que se manifestaran tarde o temprano diferentes clases de trastorno mental. Esta persepectiva se acompañaba de la idea de que las razas podían ser clasificadas en función de su supuesto grado de progreso evolutivo, con lo que algunas (en particular la caucásica) se consideraban más avanzadas tanto cultural como biológicamente que otras. El hecho de que los individuos con determinados tipos de deficiencia mental (síndrome de Down) tuvieran rasgos morfológicos semejantes a los de razas diferentes a la caucásica se consideraba prueba de ello. Además de los idiotas mongólicos, Down (1866) distinguió también, a partir de sus rasgos morfológicos, entre los idiotas de la variedad etíope, los de tipo malayo y otros semejantes a los habitantes primitivos de América.
Paul Broca fue uno de los científicos de mayor prestigio que abrazó en el siglo XIX la creencia sobre la existencia de una jerarquía de desarrollo entre las diferentes razas humanas.

Gould
(1981) señala que sus métodos de cuantificación de variables eran extraordinariamente
rigurosos, pero que adolecía de ciertos defectos a la hora de elaborar conclusiones con
los datos que recogía. Era propenso, por ejemplo, a escoger aquellos datos que ilustraban
prejuicios previos, ignorando los que los contradecían. Entre las características
morfológicas que intentó establecer para fundamentar su afirmación respecto a la
jerarquía existente entre las razas se encontraba la proporción entre la longitud del
radio y la del húmero; una mayor proporción indicaría un mayor tamaño relativo del
antebrazo, algo característico de los monos. En los individuos de raza negra la
proporción era de 794, mientras que en los blancos era de 739; no obstante, en los
esquimales era de 703 y en los aborígenes australianos era de 709, así que concluyó que
la elongación del antebrazo no era un indicador adecuado del grado de desarrollo racial
(Gould, 1981). Otros intentos de Broca para encontrar indicadores de clasificación
jerárquica racial, igualmente fallidos, fueron el ángulo facial (la proyección anterior
del rostro y la mandíbula), el índice craneano (la proporción entre el ancho máximo y
el largo máximo del cráneo), la situación del foramen magnum (agujero situado en la
base del cráneo, por el que pasa la médula espinal; los pueblos inferiores debían tener
un foramen magnum situado más atrás, como en los monos), y la proporción de materia en
las zonas anteriores del cerebro respecto a las posteriores.
La hipótesis de la degeneración contó en psicología con defensores tan relevantes como H.H. Goddard (Andrés, 1989), estudioso de la deficiencia mental e introductor de los tests de Binet en EE.UU.

Goddard participó en el programa de selección y control de la inmigración masiva que tenía lugar en EE.UU. a principios del siglo XX, aplicaba para ello pruebas de inteligencia y es conocida la caracterización como débiles mentales que hacía de marginados, delincuentes e individuos de clases bajas, explicando su situación social por su escasa inteligencia (Gould, 1986). De manera similar, Gámbara escribía en España, en un manuscrito no fechado:
"La antropología criminal, fundada por Lombroso (...), sanciona que el delincuente, con tal que no sea de ocasión, representa siempre las razas inferiores de la humanidad, á las que, por desgracia, pertenecen los deficientes".
La antropología criminal de Lombroso también surge en el contexto de las tesis degeneracionistas. Se basaba en la idea de que los criminales tienen una peculiar organización nerviosa, de tipo degenerativo, que debía poder ser identificada a partir del estudio de su morfología corporal. Ello permitiría discriminar a los criminales innatos, facilitando con ello la objetividad de las decisiones judiciales. La aplicación de las técnicas desarrolladas para la medición de los rasgos morfológicos no se limitó al campo de la criminología, extendiéndose con rapidez a otros grupos de población marginados como los enfermos mentales.
Lombroso identificaba la criminalidad como una característica propia de especies inferiores y de los pueblos primitivos; observó, por ejemplo, que, como los delincuentes occidentales, los miembros de la tribu egipcia dinka también acostumbramban a adornar sus cuerpos con tatuajes, tenían un alto umbral de dolor y su morfología facial era semajante a la de los monos (Andrés, 1989).
Las tesis lombrosianas coincidían con el ambiente recapitulacionista que se respiraba por la época, dado que incluían en una misma categoría a los asesinos natos, los animales, las razas inferiores y los niños. Para Lombroso en los niños podían observarse algunas características propias de los criminales como las tendencias sádicas y la pasión por las bebidas alcohólicas (Lombroso, 1895, p. 56).
Algunas de esas ideas degeneracionistas y sus desarrollos posteriores siguen vivas en la actualidad, como la de la estrecha relación entre psicopatía y psicopatología sobre la que se fundamenta la dimensión de psicoticismo de Eysenck; o la de la adaptación de la pena a la personalidad y las circunstancias del delincuente. Ejemplo de ello son las reformas legales basadas en el principio de la indeterminación de la sentencia y en la atención a las circunstancias atenuantes (siguiendo un modelo lombrosiano ampliado que incluye los factores ambientales junto a los personales entre los determinantes del delito), obligando a erradicar la aplicación automática de la pena en función del delito cometido como defendía la criminología clásica. La pena, de acuerdo con estas tesis, no debería ser un castigo del crimen, sino una defensa de la sociedad ajustada a las características de la persona que ha cometido el delito. No tiene sentido castigar severamente a una persona que comete un delito ocasional presionado por circunstancias ambientales; en cambio, conviene castigar con dureza al delincuente nato aunque su falta haya sido menor puesto que la sociedad ha de protegerse de él, debe segregarle.
La tesis de la degeneración fue también utilizada por algunos psiquiatras franceses durante la segunda mitad del siglo XIX para explicar, además de la locura o la criminalidad, la tendencia a la agitación política en las clases bajas. Se argumentaba que la revolución es promovida por individuos predispuestos genéticamente al alboroto. De esa manera, los miembros de clases bajas que promovían conflictos sociales eran calificados como incapaces de pensar racionalmente y carentes de libre albedrío. Los problemas sociales se atribuían a desigualdades naturales entre razas y clases. Dowbiggin (1985) sugiere que muchos médicos adoptaron las tesis degeneracionistas porque se encontraban en un ambiente en el que sufrían una extraordinaria presión por parte de la sociedad. Se tenía una baja consideración hacia los médicos que se ocupaban de la atención a las personas con enfermedades mentales, dado el escaso éxito que obtenían en sus tratamientos. Si bien Esquirol había considerado a la predisposición hereditaria una de las muchas causas posibles de la locura, entre las que citaba también a factores ambientales igualmente importantes como los estilos de vida, el clima, etc, otros autores como Moreau se decantaban claramente por la transmisión hereditaria de la tendencia a la degeneración como causa principal. La patología mental podía ser inicialmente consecuencia de condiciones sociales o ambientales, pero una vez adquirida se transmitía como una degeneración hereditaria. No se heredaría una enfermedad determinada, sino una tendencia a la degeneración que podría expresarse en forma de diferentes enfermedades.
 |
El origen de los problemas mentales, de la
criminalidad y, en general, de cualquier tipo de desviación era atribuido a factores
hereditarios. En este contexto, Sir Francis Galton dió el nombre de eugenesia
a una disciplina que tendría como finalidad sustituir la selección natural por una
selección social encaminada a evitar la degeneración de nuestra especie. Eran los
primeros pasos del darwinismo social. Hubo notables defensores de la eugenesia, desde que
Galton formulara sus objetivos en 1883; entre ellos, ya entrado el siglo XX, se
encontraron biólogos como Davenport y psicólogos como Burt y Terman. Para alcanzar sus
fines la eugenesia precisaba poner énfasis en la medición cuantitativa de los rasgos
físicos y psicológicos, lo que hizo que simpatizaran con ella muchos psicólogos
embarcados en la tarea de la construcción de tests mentales. Esta tradicional
vinculación de los tests de inteligencia con la eugenesia hizo que aparecieran numerosas
críticas ideológicas a aquellos, y muchas de ellas se mantienen en la actualidad
(Andrés Pueyo, 1997) La eugenesia y el darwinismo social fueron abrazados en los años 30 de nuestro siglo como respaldo "científico" de los movimientos racistas y fascistas. Ya en 1911 el estado de Nueva Jersey había promulgado una ley estipulando la esterilización de retrasados o enfermos mentales, criminales, borrachos, drogadictos, epilépticos, sifilíticos y otras personas "enfermas y degeneradas" (Chorover, 1979). |
Información complementaria: Etología y sociobiología
En consonancia con la visión spenceriana de la evolución, Jackson (1955) propuso el concepto de encefalización, con el que se asumía que las regiones nerviosas de aparición más reciente dominaban la actividad de las precedentes. Su idea principal era que la evolución del sistema nervioso central (SNC) se había caracterizado por la adición sucesiva de estructuras; cada nueva estructura servía para inhibir selectivamente y, por ello, para incrementar la selectividad de respuesta de las estructuras inferiores. Las estructuras inferiores eran automáticas mientras que las superiores evidenciaban la participación cada vez mayor de la voluntad en el control del comportamiento. Cuando una estructura superior como el córtex era dañada, se desinhibía la función normal de la estructura inferior; se producía una regresión. Por tanto, el comportamiento controlado por centros nerviosos inferiores era, según Jackson (1955), resultado de la disolución de los centros superiores, y ésta podía ser producida por causas como las lesiones cerebrales, la vejez o la enfermedad. Esta idea establecía, como se ha comentado anteriormente, un cierto paralelismo entre los trastornos neurológicos, la enfermedad mental, la deficiencia mental, el comportamiento de los llamados pueblos primitivos, el de los criminales y el de los organismos inferiores en la escala evolutiva; así Goddard (1919) escribía:
"La inteligencia controla las emociones y las emociones están controladas según el grado de inteligencia que se tenga (...), cuando la inteligencia es pequeña, las emociones no están controladas (...) se traducirán en actos desordenados, descontrolados y generalmente delictivos".
Una versión contemporanea de los supuestos jacksonianos es representada por McLean (1993), para quien es posible distinguir tres componentes evolutivos en el cerebro humano: la parte más antigua, ya presente en los reptiles, sobre la que se añadiría el cerebro límbico propio de los paleomamíferos y, finalmente, la corteza cerebral propia de los neomamíferos; correspondiendo a cada uno de estos tres componentes funciones de control del comportamiento progresivamente superiores: automáticas, emocionales y cognitivas.
Como en Jackson, en McLean se encuentra el presupuesto implícito de que el proceso evolutivo de las especies ha sido unidireccional, con cambios sucesivos hacia el patrón humano, considerado como el más evolucionado. Este presupuesto (o los dos que conlleva, es decir, direccionalidad y superioridad) coincide con la visión que tenía Spencer de la evolución, pero es ajeno a la de Darwin, para quien la evolución ha seguido diferentes líneas que dan lugar a una gran diversidad de formas de vida, cada una de ellas adaptada de una manera concreta (pero no mejor ni peor que otras) a su entorno (Mayr, 1991; Ruse, 1986; Armstrong, 1990; Dubrovsky, 1993).
 Información
complementaria: Brain Evolution and Development. The Selection of Neurons and Synapses
Información
complementaria: Brain Evolution and Development. The Selection of Neurons and Synapses
Tales ideas han influido considerablemente sobre la neurología, la psicología y la psicopatología desde entonces. Freud (1966), por ejemplo, propuso que el Yo y el Super-yo iban adquiriendo gradualmente control sobre una instancia biológicamente más primitiva como el Ello y, al igual que Janet (1903), explicó la enfermedad mental como un fracaso en el control que las instancias superiores ejercen sobre las inferiores. Expresiones más recientes de esta forma de pensar corresponden a Ey (1969), para quien "ser demente, oligofrénico, esquizofrénico, maníaco, melancólico o neurótico es permanecer o regresar a un nivel inferior de organización de la vida psíquica".
Investigaciones fisiológicas realizados por autores de gran prestigio como Hubel y Wiesel (1965, 1968) han contribuido en años recientes a asentar firmemente la idea de jerarquía. Diferentes estudios como los de Zeki (1974), Goldman-Rakic (1984), y Zeki y Ship (1988), no obstante, hacen pensar que, si bien el concepto de jerarquía sigue siendo útil en la actualidad, éste no debe referirse a una sola cadena jerárquica, sino a múltiples vias paralelas relacionadas con el procesamiento de distintos atributos del estímulo y que constan, cada una de ellas, de una cadena jerárquica diferenciada (Dubrovsky, 1988, 1993; Zeki, 1995).
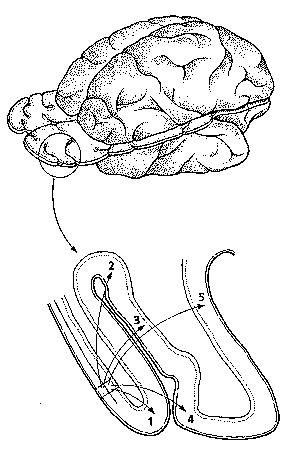
En este punto coinciden tanto los neurobiólogos experimentales como los neurobiólogos computacionales, puesto que desde ambas perspectivas el modelo resultante de organización del sistema nervioso pasa a ser el de un conjunto de subsistemas distribuidos de forma paralela. Cada área cortical tiene múltiples salidas; ello hace que los resultados de las operaciones realizadas en una región determinada se transmitan a otras muchas áreas, sin que ello signifique que se transmita la misma operación a todas ellas, puesto que cualquier región puede emprender varias operaciones de manera simultánea, gracias al agrupamiento de las células con propiedades comunes (Goldman-Rakic, 1984), y distribuir los resultados respectivos a varias zonas corticales distintas (Zeki, 1974, 1995). Los modelos jerárquicos secuenciales van siendo, por tanto, sustituidos por modelos basados en los conceptos de paralelismo, segregación-especialización funcional, y multiplicidad de áreas en cada modalidad sensorial.
El esquema jerárquico de la organización del sistema nervioso propuesto por Jackson se relaciona estrechamente con la noción de que los procesos sensoriales y perceptivos son secuenciales (Dubrovsky, 1988). Según esta idea, el reconocimiento y la identificación se llevan a cabo a través de una serie de etapas en diferentes niveles de complejidad. Por ejemplo, los estímulos visuales son procesados inicialmente en la retina, a continuación en el tálamo, luego en la corteza visual primaria y, por último, en las áreas corticales de asociación, donde tenía lugar la percepción consciente a través de la integración de las señales sensoriales procedentes de áreas primarias adyacentes. De acuerdo con este modelo jerárquico secuencial, se pensaba que las areas de asociación no recibían proyecciones talámicas (Campbell, 1905). Sin embargo, algunos estudios comentados por Rose y Woolsey (Dubrovsky, 1988) demostraron que las zonas de asociación también reciben tales aferencias; por otro lado, los estudios de Diamond (1983) con el mamífero insectívoro tupaia glis mostraron que la resección de la corteza estriada no impedía moverse por el entorno, evitar obstáculos, alcanzar el alimento, seguir objetos en movimiento y aprender hábitos basados en discriminaciones visuales; cuando las lesiones se extendieron a areas adyacentes a las estriadas, los tupaia sufrieron una pérdida acentuada de las discriminaciones visuales aprendidas. Estos resultados confirmaban que las zonas denominadas de asociación recibían señales que llegaban en paralelo desde el tálamo; es decir, no dependían en su totalidad de aferentes corticales provenientes de las areas corticales primarias.
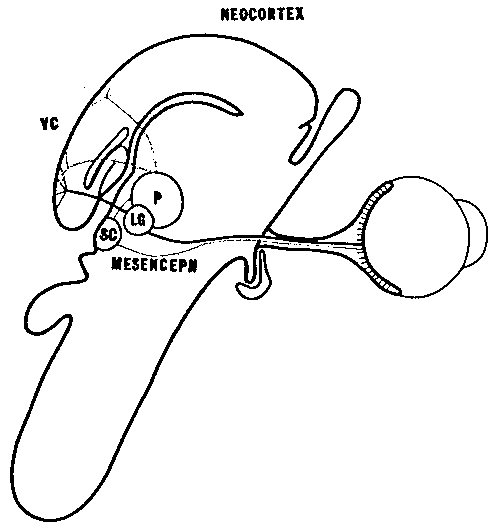
A partir de éstos y otros estudios relacionados con el procesamiento de otros tipos de señales, como las hormonales, Dubrovsky (1988), plantea la hipótesis de que el procesamiento que el sistema nervioso hace de las señales no es característicamente secuencial (como se derivaría de una organización jerárquica) sino en paralelo. En realidad el procesamiento en paralelo no elimina sino que extiende el procesamiento en serie.
En psicopatología las propuestas de Jackson se han traducido en la presuposición de que existe un continuo entre la normalidad y el trastorno mental. En palabras de Jackson (1955): "la enfermedad no crea, libera". Las funciones normales pueden así ser extrapoladas a partir de las funciones patológicas, y viceversa. Esta concepción de las relacionaes entre la normalidad y la anormalidad es consistente con la tradición galénica que entendía la enfermedad y la personalidad como el resultado de un balance determinado de los humores corporales, que cuando están en equilibrio dan lugar a la personalidad normal. Una de las expresiones actuales de la doctrina de los humores es la investigación basada en el establecimiento de las relaciones entre el trastorno mental y el déficit o el exceso de determinados neurotransmisores. Se ha propuesto que el mecanismo de acción de algunos psicofármacos consiste en el incremento (p.ej. los antidepresivos tricíclicos o los inhibidores de la monoaminooxidasa) o la disminución (p.ej. los neurolépticos) de la disponibilidad de neurotransmisores en la sinapsis, lo que ha contribuido en gran medida a respaldar la hipótesis de la continuidad entre la normalidad y el trastorno mental. Esta idea, sin embargo, no ha dejado de ser objeto de discusión en la actualidad; Dubrovsky (1990), por ejemplo, sugiere que el trastorno mental no puede ser entendido exclusivamente como una pérdida del balance de neurotransmisores, sino que ha de ser abordado como un estado cualitativamente diferente de la normalidad, resultado del funcionamiento del sistema nervioso de un modo diferente al normal, bajo cuyas condiciones no son aplicables sin modificaciones significativas las leyes de organización neural propias de la normalidad.
En apoyo de la hipótesis de la discontinuidad pueden citarse algunos estudios realizados con pacientes fóbicos. Una de las características clínicas de los sujetos con fobia es el temor anticipatorio ante determinados objetos, por lo que Dubrovsky, Barbas y Solyom (1978) platearon que el paradigma de la variación contingente negativa (CNV) podía ser apropiado para estudiar la actividad cerebral de estos pacientes. La CNV es una onda que refleja un potencial negativo lento dependiente de la asociación de dos estímulos sucesivos. En el paradigma experimental básico un estímulo, S1 (también denominado estímulo condicional o de preparación), es seguido tras un intervalo fijo de tiempo por otro estímulo, S2 (también denominado estímulo incondicional o imperativo), que es señal para el sujeto dé una respuesta (como apretar un botón). La CNV se produce en dos fases, una primera de orientación ante S1 y una segunda de expectativa ante S2. Cuando S2 es una reproducción del estímulo fóbico, los registros CNV de los pacientes muestran mayor amplitud que cuando el apradigma no contiene el estímulo fóbico, así como una gran semejanza con los que se encuentran en algunos pacientes psicóticos en los que aparece una prolongación de la onda denominada variación negativa postimperativa (PINV). Previamente, Knott e Irwin (1973) habían utilizado un paradigma similar (anticipación de un estímulo temido, una descarga eléctrica) con personas normales, informando de registros CNV con amplitud más reducida y resolución más rápida de la onda negativa que la que posteriormente encontraron Dubrovsky et al. (1978) en sujetos con fobia específica. En este mismo artículo se informa que tras un tratamiento con desensibilización sistemática, tanto las manifestaciones clínicas como las características de la CNV de los pacientes retornaron a la normalidad. Como explicación de tales resultados propusieron que el emparejamiento de una condición placentera, como la relajación, con el estímulo inicialmente fobogénico, establece nuevas asociaciones a través de las estructuras límbicas implicadas en los procesos emocionales. Las conexiones entre el sistema límbico y los lóbulos frontales pueden actuar entonces como moduladoras de las regiones frontales desde las que se genera, principalmente, la CNV. Para este modelo la fobia no es, por tanto, un miedo muy intenso, es decir, una extensión de un fenómeno que existe normalmente, sino una nueva clase de respuesta; lo que hace que un estímulo se convierta en fobogénico es que, además de la activación de las redes neuronales que provoca en condiciones normales, ahora elicita respuestas de neuronas que normalmente no reaccionan ante él (Dubrovsky, 1993). De manera general se puede decir que el concurso de diferentes factores físicos, químicos y ambientales puede llevar a una red neuronal concreta a colocarse en un "modo" determinado de funcionamiento (Bullock, 1993); es decir, una tendencia a procesar la información o generar respuestas de determinada manera. Cada modo representa una organización de la red que da lugar a una función (Getting, 1989). La transición entre modos está determinada por las aferencias que se reciben o por cambios en el sustrato químico que alteran las propiedades de los componentes elementales de la red (Dubrovsky, Filipini, Gijsbers y Birmingham, 1994). Por ejemplo, las entradas aferentes a determinadas redes neuronales no sólo sirven para activarlas, también las configuran en uno de diferentes circuitos funcionales posibles, como se ha comprobado en la tritonia (Getting, 1989).