|
U n i v e r s i d a d d e B a r c e l o n a |
Departamento de Personalidad, Evaluación y Tratamientos Psicológicos |
Cursos asistidos por ordenador a través de i n t e r n
e t |
|
U n i v e r s i d a d d e B a r c e l o n a |
Departamento de Personalidad, Evaluación y Tratamientos Psicológicos |
Cursos asistidos por ordenador a través de i n t e r n
e t |
| Neurotransmission
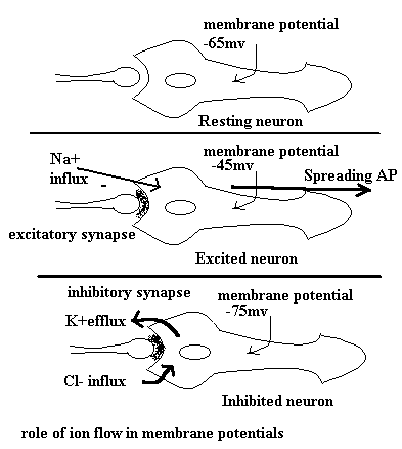
Synaptic biologyThe Action Potential and Synaptic Functions The method that the neurons use to process information at the cellular and network level is based on the action potential. The action potential is in turn an expression of the structural components of the neuron. The neuron has four basic structural elements which process information. These are the body (soma), dendrites, axon, synaptic endings. The essential element of the neuron that allows neurons to communicate with each other is the electrical characteristics of the membrane. The membrane is a semipermeable lipid bilayer. The electrical potential of the membrane is the result of the selective permeability of the membrane to ions. This selectivity is the result of "gates and pumps." A gate is a 'pore' that allows ions to flow down an electrochemical gradient (i.e., from a high concentration to a lower concentration). A 'pump' is an enzyme related complex that pulls an ion against an electrochemical gradient. The best know pump is Na/K ATPase. The pumps are energy dependent and probably account for most of the energy utilization in the CNS. In an undisturbed system the gradients settle at the resting membrane potential. The potential can be calculated by knowing the intracellular and extracellular concentrations. Huxley and Hodgkin received the Noble Prize for their description of the membrane electrical gradients. Membrane events related to synaptic interactions cause alterations
in the ion gradients. If the interaction allows cations such as Na+, K+, or Ca++
to flow into the cell more easily a depolarization occurs. If the interaction allows Cl-
ions to flow into the cell more easily then a hyperpolarization occurs. If the sum of
these interactions reach a threshold then the cell generates a large stereotypic response
at the junction of the axon and the soma. This response is a self propagating action
potential that depolarizes down the length of the axon without fail. The action
potential is the result of several changes in the conductance of the ions. The changes are
the result of voltage gated sodium and potassium pores. The figure below illustrates the
voltage gating process. At the end of the axon the action potential reaches the synaptic
terminals. The events in the synaptic terminals result in discharge of neurotransmitters
to a second cell. The rate at which the action potential travels down the axon is
increased by the presence of myelin. We will discuss the formation of myelin and
myelinopathies in a future lecture.
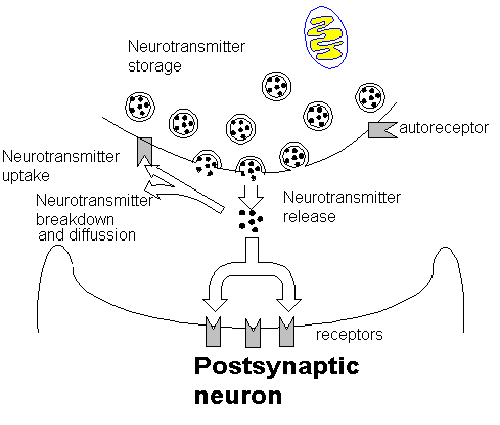
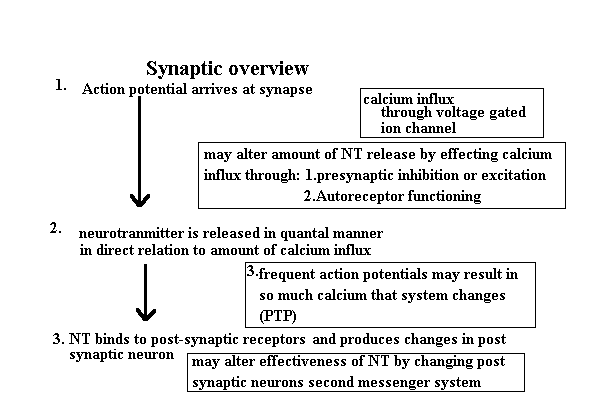
Synaptic Biology: General Principles Neurons process information through the synaptic connection of the brain. The structural features of the synapse are intimately related to biology of the synapse. Although there are differences among the synapses of the various neurotransmitters the general principles will be described in this section. There are two basic types of neurotransmitters; small molecular weight and peptides. The small molecular weight neurotransmitters include Acetylcholine, monoamines, and amino acids. The peptide neurotransmitters include opiates, and other small peptides. An individual neuron may have a small molecular weight neurotransmitter co-localized with a peptide neurotransmitter. The regulation of the synthesis of these two neurotransmitter types varies. The peptides are synthesized in the soma and during transport down the axon may undergo post translational modification. Thus the regulation of the synthesis of the peptides involves the genetic apparatus primarily. The small molecular weight neurotransmitters are synthesized primarily in the synaptic terminal and the regulation of synthesis is affected acutely by changes in the presynaptic terminal. This allows the regulation of the small molecular weight neurotransmitters to be more responsive to rapid changes. This also means that changes in the peptide neurotransmitter's rate of synthesis and metabolism are more enduring changes. This difference in fundamental regulation of the peptides and small molecular weight neurotransmitters suggests a difference in the roles they play in neuronal communication. The small molecular weight neurotransmitters are more point to point communications while peptides may exert a more long term effect on post synaptic neurons. The functional components of the synaptic system are basically the pre and post synaptic neurons and the synaptic cleft. The simplest synaptic unit is conceptualized as a two neuron, one transmitter system. However, this situation can be come rapidly more complex as the number of cells and neurotransmitters and types of synapses increase. The diagram below describes the events leading to the cell surface interaction of the neurotransmitter and the post synaptic cell.
The neurotransmitter 'package' is released into the synapse through a process of exocytosis. Exocytosis is calcium dependent. Normally calcium is present in the intracellular fluid bound to proteins so that it is inactive. The calcium that powers the exocytosis is primarily the result of the calcium influx through voltage dependent gates. These gates increase the conductance to calcium when the action potential arrives. The amount of calcium that enters the terminal is dependent upon how 'wide' these gates open. The gates open in direct relationship to the difference between the resting membrane potential at the time the action potential arrives and the peak of the action potential. If the membrane potential is hyperpolarized prior to the action potential's arrival the difference is greater and more calcium enters and more exocytosis occurs and more neurotransmitter effect results. If the presynaptic membrane is partially depolarized prior to the arrival of the action potential then there is less difference between the membrane potential and the peak of the action potential and hence less calcium influx and less neurotransmitter effect. Such effects on the membrane potential occurs through presynaptic inhibition and facilitation. The neurotransmitter release can also be influenced by a process known
as potentiation. In this situation the neuron has had repeated action potentials arriving
at the synaptic terminal. The result is a greater and greater amount of calcium in the
synaptic terminal. The calcium soon overwhelms the available binding sites and with each
action potential more calcium is free to power the exocytosis. Therefore there is more
neurotransmitter effect. This potentiation is sometimes referred to as post tetanic
potentiation. It is a form of early molecular memory according to some sources. If the
activity continues at a high level then the neuron begins to develop a reduced release of
neurotransmitter in a process known as habituation. Regulation of the amount of
neurotransmitter released can be affected by altering the amount of calcium in the
synaptic terminal or by altering the degree of voltage change which occurs as the action
potential arrives at the synapse.
Once the neurotransmitter is released there are three basic things that
can happen to it. It can diffuse out of the cleft, it can be taken back up by
the presynaptic neuron, or it can bind to a receptor on the post synaptic neuron.
The rate of binding depends on the affinity of the receptor for the molecule and the
actual neurotransmitter and receptor concentrations. Once the neurotransmitter and
receptor combine there is a change in the behavior of the post-synaptic neuron. The
changes are typically the result of an alteration in either the ion channels or second
messenger systems. Of the neurotransmitter-receptor interactions known only a few
actually result in direct effects on the ion channels. These receptors are known as ligand-gated
channels or ionophore complexes. The best example in psychiatry is the
GABA-benzodiazepine-chloride ionophore complex. The other neurotransmitter-receptor
interactions are mediated by second messenger systems. These neurotransmitter-receptor
systems are termed G-protein related systems. The ligand gated channels respond
very rapidly while the G-protein systems are a bit slower in response. Receptor subtypes
are believed to be consistent in whether they are ligand gated or G-protein related. The
catecholamines (dopamine and norepinephrine) receptors are G protein related. Ligand
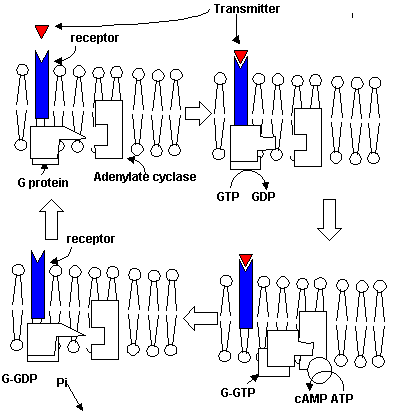
gated channels are typical of GABA, glutamate, and a few others. Signal transduction Transmitter-receptor interactions are extracellular for the most part. The goal of synaptic processes is to cause an effect on the intracellular processes of the post-synaptic neuron. This is accomplished through a process known as transduction. It is at this point that the G protein is probably the star. When the transmitter couples to the receptor, a conformation change occurs in the receptor. This conformational change results in an increased affinity for the G-protein which is on the inside surface of the cell membrane. The G-protein can then act to either couple the receptor-transmitter complex to an ion channel, or regulate the synthesis of second messengers.
The effects of G-protein on second messenger systems are dependent on whether the G-protein is G-s (stimulating) or G-i (inhibiting). The effect of most second messenger systems is on protein phosphorylation rates. The best understood second messengers are cAMP, phosphatidyl inositol, cGMP, and calcium. Nitric oxide and the polyamines are less well understood and considered second messengers by some authors. A discussion of the cAMP and phosphatidyl inositol system is in order. cAMP is produced by adenylate cyclase in response to G-s protein/receptor interactions. (G-i would cause an inhibition of adenylate cyclase.) cAMP activates a cascade of enzymatic processes through a cAMP dependent protein kinase. This enzyme phosphorylates a number of proteins with wide ranges of effects. cAMP has a widely distributed effect on almost all cellular processes including the basic genetic transcription via the CREs described in the next section. cGMP is basically analogous but differs in the range of substrates for the cGMP dependent protein kinase. We have discussed the regulation and effects of calcium above.
Calcium is the only ion that qualifies as a second messenger. This is due to the
fact that calcium is capable of binding to regulation sites on a number of enzymes
including Protein Kinase C and a Ca/calmodulin dependent protein kinase. These kinases
yield similar cascades as cAMP and cGMP through phosphorylation of substrates.
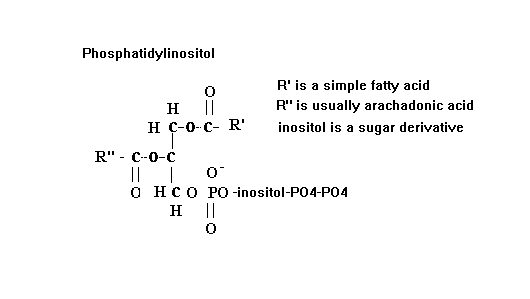
The final second messenger we will describe is the phosphatidylinositol system. Recall that phosphatidylinositol is a phospholipid that is present in the cell membrane. The general formula is shown below.
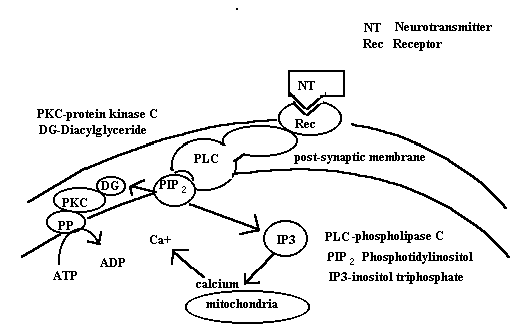
Neurotransmitter-receptor interactions result in the activation of
phospholipase C that cleaves the phosphoinositol from the third carbon in the glycerol
backbone of the molecule. The result is the formation of diacylglycerol and inositol
triphosphate. DAG and IP3 are the active molecules in the cascade illustrated below. IP3
displaces calcium from intracellular stores that then activates calcium dependent kinases.
DAG activates protein kinase C which phosphorylates enzyme substrates as in the other
second messenger systems.
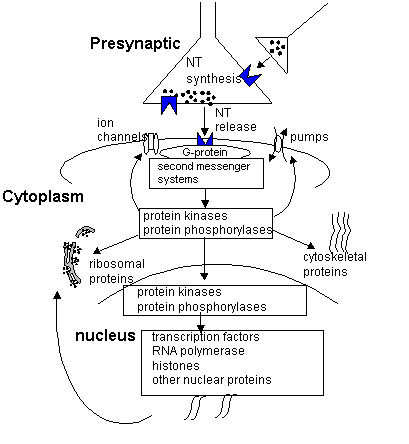
Molecular Biology and genetics The information flow across neurons does not end at the second messenger level. . The cell machinery of the neuron is specialized in order to facilitate the information processing. The most important components of the intracellular system are the genes. The DNA which comprises the genes is an extended molecule capable of responding to the flow of information in the individual neuron. The genetic code of the neuron, codes for the amino acid sequences of the cellular machinery. Information flow from the nucleus and the gene to the final protein product is complicated but sheds some insight into learning, memory, and the response of the nervous system to stress. The fact that behavioral responses changes during treatment with psychotropic agents suggest that there is a dynamic regulation of the expression of genetic information in the neuron. Therefore a review of basic molecular biology is relevant in the discussion of psychopharmacology.
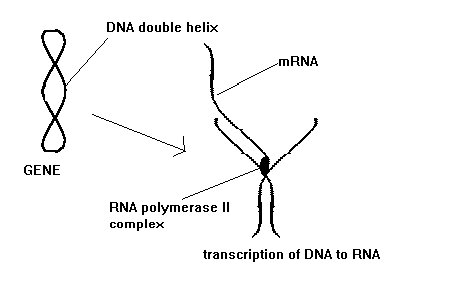
Recall that the information of the DNA is transcribed into mRNA in the nucleus. The messenger RNA is then translated into protein outside the nucleus. Transcription occurs in three steps: initiation, elongation, and termination. Regulation of the transcription can be accomplished at any of these steps. In the nervous system the principle site of regulation is the initiation step. In eukaryotes the transcription step is carried out by a complex of proteins that includes RNA polymerase II. The polymerase II complex regulates transcription . For it to initiate transcription the complex must be positioned at a start site of a gene. The rate at which this complex produces transcription mRNA is controlled by factors known as transcription factors. There are literally hundreds of these proteins that act as activators or repressors of gene transcription. The genes for these factors are not necessarily close to the genes they activate. The section of genes that they activate can be thought of as receptors for the transcription factors. The expression of the transcription factors and the gene receptor areas are controlled by the cell type, developmental stage, and even specific physiologic stimuli. As the process of the sculpting of the circuitry of the brain occurs certain pathways are strengthened and certain pathways are lost. This activity-sensitive process is due to the ability of the synaptic events to alter gene expression. Since all cells have the same DNA there must be a differential expression of genes. This differential expression is the result of three mechanisms. The first includes the cell-type specific elements of the DNA binding sites. These sites are expressed as a result of an interaction in specific cell types. The second mechanism is the result of "response elements." These elements mediate gene expression in response to a particular physiologic stimulus. Glucocorticoids are an example of a response element. Other hormones such as gonadal, thyroid, and vitamin D act as response elements in the nervous system. These hormones are bound to cytosolic receptors that are then translocated into the nucleus where they bind to the DNA to either augment or repress transcription activity. The third mechanism of differential gene expression is probably the most important and is the result of second messenger response elements. The best understood example of this is the Cyclic AMP response element (CRE). cAMP activation leads to the binding of protein transcription factors (CREBs) to the DNA at sites termed CREs. The binding of the CREBs is influenced by phosphorylation by cAMP-dependent protein kinase. Analogous to the cAMP system is a system that confers activation in response to protein kinase C signal transduction pathway. The protein transcription factors include the AP-1 complex. The best example of this complex is the c-FOS and c-JUN oncogens. The remarkable aspect of this transcription system is the ability
for the environment to mold and shape the processing pathways. Such learning is
accomplished at a molecular level. The signal processing pathways can be facilitated by
cell specific effects on transcription, hormonal effects and second messenger system. This
explains the dynamic regulation of the expression of genetic products in neural networks.
Translation of mRNA into proteins The transcription of DNA into RNA complement forms a strand of mRNA. The mRNA is translocated out of the nucleus into the cytoplasm where the code is read. Recall that the code is composed of triplet bases which each code for a different amino acid. The triplets on the mRNA are referred to as the codon. Transfer RNA has a sequence that has the complementary triplet known as the anticodon. At the 3' end of the tRNA the amino acid is located. At the ribosome the tRNAs loaded with amino acids locate the codon specific for their anticodons. This causes the growth of a polypeptide in a predetermined sequence. At the end of the mRNA the ribosome separates into subunits and releases the polypeptide. The newly translated peptide may be further modified through a process of post translational modification. The post translational modification process is another point at which the cellular machinery can be regulated. In general there are two basic ways to modify the activity of a protein. The first is by allosteric modification. A noncovalent interaction with another molecule leads to a conformation change which then leads to a change in function. This is a common site of enzyme regulation in metabolic pathways. The second type of modifications are covalent modifications whereby the protein is altered by the addition of a sugar group (glycosylation), phosphate group (phosphorylation), or acyl groups (acylation). In addition covalent modification, structural changes can be accomplished by formation of disulfide bonds to hold a conformation and cleaving specific sequences to expose the active sites of the molecule. The newly synthesized protein can be cleaved to yield several daughter
products. This is done in the pro-opiomelanocortin molecule. These daughter products
allows the cell to create several products from the same mRNA. Neurotransmitter SystemsCatecholamines; synthesis and storage The catecholamines that act as neurotransmitters include dopamine and
norepinephrine. Epinephrine is not produced in the central nervous system. The
catecholamines share a common synthetic pathway. The rate limiting step is tyrosine
hydroxylase. The activity of tyrosine hydroxylase can be modified by phosphorylation. This
provides a point of regulation for the neuron. Clinically there are no agents capable of
modulating tyrosine hydroxylase. However, treatment with agents such as methyldopa can
compete with dopa for further processing. The result is a formation of false
neurotransmitters. The false neurotransmitters are packaged in the synapse as though they
were the catecholamine but when released into the synapse they are ineffective at the
receptor. False neurotransmitters such as octopamine are also thought to be increased in
hepatic encephalopathy. The packaging of false transmitters in the periphery is thought to
be increased by inhibiting MAO. This is thought to explain the orthostatic blood pressure
effects caused by therapeutic doses of MAO inhibitors.
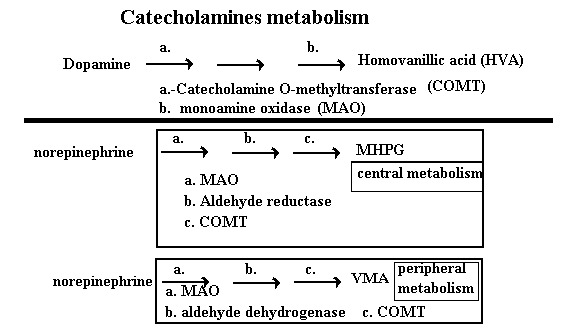
The major biochemical difference between the norepinephrine and dopamine neurons is the presence of a vesicle bound copper containing enzyme in the neurons of the norepinephrine neurons. This enzyme is dopamine beta hydroxylase. It catalyses the synthesis of norepinephrine from dopamine. The catecholamines have similar metabolic pathways. The chart below shows these pathways. A point of potential therapeutic intervention in metabolism is at the level of monoamine oxidase. MAO is primarily found associated with the mitochondria. The intracellular site of MAO suggests it is involved in metabolizing catecholamines once they have been taken back up into the neuron to prevent repackaging them into the synaptic vesicles. There are two MAO isoenzymes. MAO-A appears to be a specific enzyme for serotonin and norepinephrine. MAO-B is a more broadly active enzyme that acts on phenylethylamines. Tyramine and dopamine are equally good substrates for both MAO-A and MAO-B. There is an anatomic distribution of the isoenzymes. MAO B is found primarily in the brain. MAO A is found thought the body and especially in the GI tract where it may serve to limit the absorption and physiologic effect of dietary monoamines. There are specific inhibitors of each of these. Nonspecific MAO inhibitors must inhibit all the MAO. Because specific MAO inhibitors only inhibit one of the isoenzymes there is always MAO activity present. This has a couple of consequences; first there is less chance for a tyramine effect, second there is less orthostatic effect.
As you see from the chart the catecholamines are also metabolized by catechol-O-methyl
transferase (COMT). COMT is present extracellularly. MAO can act before or
after COMT of the catecholamines. Make note of the metabolic products as these can reflect
the turnover of the neurotransmitters. Catecholamine synapse The norepinephrine and dopamine synapses are basically analogous.
Important extracellular features include the presence of reuptake sites, multiple post
synaptic receptors, and autoreceptors and presynaptic receptor sites. The 'function' of a
neurotransmitter depends on a combination of the network it is being used in, and the
post-synaptic receptor's behaviors. The autoreceptors modulate the release of
neurotransmitter by the presynaptic neuron. The autoreceptors are believed to have a
difference in affinity with the neurotransmitter. This is shown by the fact that extremely
low doses of a receptor agonist are inhibitor to the release of endogenous transmitter
and low dose of a receptor antagonist augments release of neurotransmitter. It is believed
that this explains the early increase in HVA that occurs when neuroleptics are first
initiated. Some neurons appear to have both presynaptic facilatory and inhibitory
receptors. The presence of both a presynaptic facilitating and presynaptic inhibitory
autoreceptor suggest the need for fine tuning of the release of the neurotransmitter. It
should be obvious that the relative proportion of each can adjust the set point of the
neurons release of neurotransmitter much like the max and mins of a thermostat adjust the
activity of a furnace in the home. Enduring changes in activity can be acheived by
altering the genetic expression of one or both of the genes for these autoreceptors. In
general the mechanism of the effects of these presynaptic autoreceptors are the same as in
presynaptic inhibition or facilitation. That is they modulate Ca++ channels or influence
the membrane potential of the presynaptic neuron or they influence adenylate cyclase
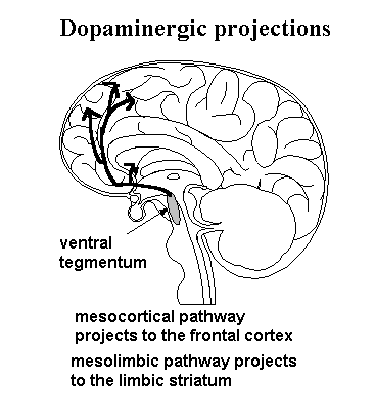
activity. Catecholamine projections The behavioral effect of a pharmacologic intervention on a specific neurotransmitter is related to a number of things. An idea of the behavioral effects of the catecholamine pathways are disclosed by considering the sites of projection. The diagrams below illustrate the projections of the dopamine systems and norepinephrine systems in the human brain. .
There are two major cell groups of norepinephrine containing cells in the neuroaxis. The First arises from the locus ceruleus in the caudal pontine grey and spreads anteriorly forming an extensive net throughout the cortex. The second cell group projects from the lateral tegmental neurons proceed caudally to the spical cord and rostrally into the diencephalon. The axons of the LC neurons have varicosities which allows a widely distributed release of norepinephrine. It is felt that this reflects a neuromodulatory function of this system. There are two norepinephrine tracts relevant to behavior which lead out of the LC and lateral tegmental neurons; the dorsal bundle and the median forebrain bundle. The widespread network of these neurons includes innervation of specific hypothalamic and thalamic cell groups. There are several theories of the role of the locus ceruleus in learning, memory, anxiety and psychosis. A more general and more understandable role can be described as a system of the orientation of the brain to stimuli in the environment and viscera. This system is activated by a variety of sensory stimuli. It seems to be related to vigilance. Such orientation response is needed to explore the environment. It is a necessary part of the central control of the autonomic nervous system. This system is also a part of the sleep-wake system as we will discuss in a subsequent section. Overactivity of the locus ceruleus system is implicated in anxiety disorders and drug withdrawal states. Another aspect of the vigalence system is its role in reward and reinforcement. It can be seen that dysfunction of the LC would result in hypothalamic dysfunction, anxiety, hedonic alterations, autonomic arousal, and sleep disturbances. It is little wonder that the LC is a focus of interest in affective disorders, anxiety disorders, and drug addiction and withdrawal. The fibers of the NE tract sweep over the anterior pole before proceeding caudally. Thus, lesions along the pathway can result in a functional decrease in NE activity. This is believed to be part of the reason that anterior strokes result in a state similar to functional depression. The fibers from the Lateral tegmental neurons proceed caudally into the spinal cord and anteriorly into the deincephalon and basal forebrain region. The basal forebrain is the region just inferior to the anterior part of the corpus callosum. This is an extremely important region behaviorally. It includes the septal nuclie which are important in the reward and reinforcement system
The projections of the dopamine system comprise five basic systems. Four of these are behaviorally relevant. These include the mesocortical, mesolimbic, tuberoinfundibular and nigro striatal systems. The mesocortical, mesolimbic, and nigrostraital systems have several similarities pharmacologically. In general the nigrostriatal system is concerned with the initiation and maintanence of motor behaviors. It runs from the substatia nigra to the caudate and putamen. There is a feedback inhibition loop which includes acetylcholine, and GABA. The relevance of this loop is apparent in situations where it is interupted. The mesolimbic and mesocortical systems arise from the ventral tegmentum. This cell group is barely separable from the substantia nigra. The mesolimbic system projects to elements of the limbic system including the amygdala, hippocampus, nucleus accumbens and the spetal area. Of significance these areas are in close proximity to the caudate and putamen. In fact some neuroanatomist call this region the limbic striatum. The mesocortical system projects to the frontal cortex. There appears to be separate feedback loops from these systems. Because of the intermingled nature of the cells of origin a feedback in the mesolimbic system also affects the mesocortical system and vis versa. The result of this situation will be considered when we discuss schizophrenia. The tuberoinfundibular tract has a major role in the regulation of some hypothalamic and pituitary peptides. Principle among these is prolactin. Inhibition of dopamine activity in this tract results in an increase in prolactin release. This explains the galactorrhea seen with neuroleptic use. Of more research interest is the potential use of prolactin to monitor central dopamine blockade. Some speculate that lack of a prolactin increase with neuroleptics predicts treatment non-response. Others have not seen such a correlation. Another aspect of this system is that it is inhibited in acute stress. This make prolactin one of the so-called stress hormones. This is the reverse of activity in the mesocortical and mesolimbic system which is activated in stress. Other important differences is the lack of autoreceptors on the presynaptic terminals inthe tuberoinfundibular system. The result is that although similar in many ways theinfundibular tract differs pharmacologically and for that reason may not be a particularly good predictor of response in the mesolimbic/mesocortical systems. The mesolimbic and mesocortical systems appear important in the initiation
and maintenance of goal directed and reward mediated behaviors. This includes the
proper maintenance of cognitive sets (ie logical thought). A dysfunction in this system
alters the normal association process and leads to a breakdown in the proper perceptual
functioning of the heteromodal
areas of the frontal lobe . This results in an inability to screen nonmeaningful
stimuli. Possible consequences would include such experiences as loosening of
associations, bradyphrenia, flight of ideas, delusional perceptions. In addition there may
also be a role of the dopamine system in regulation of affective expression. A
classic disease state of increased dopaminergic activity is Huntington's Disease.
Parkinson's disease is the classic example of dopamine deficiency. Although these diseases
are usually considered movement disorders they have parallel effects on the limbic
striatum as well. In fact these diseases are often missed when the first presentation is
to a psychiatrist. Dopamine receptor dynamics There is good evidence for at least four subtypes of postsynaptic
dopamine receptors. The D1 and D2 receptors utilize differing transduction mechanisms. D1
appears to be linked to cAMP via a G-s unit. D2 appears to be linked via a G-i unit. D2
also inhibits Ca++ entry through the voltage sensitive channels. There is also evidence
that D2 increases K+ conductance and leads to a hyperpolarized state. This would inhibit
the post synaptic neuron. Both D1 and D2 appear to have an influence on the IP3 system. A
subtype of the D2 receptor is the D4 receptor. This D4 receptor has a greater affinity
for clozapine. It is distributed in the frontal cortex, midbrain and amygdala. The
D4 receptor appears to be in low concentrations in the motor striatum. This may help
explain the general lack of EPS with clozapine. Reward (optional for now) The neurobiologic processes of reward and reinforcement influence motivational behaviors. Most of what we know comes from self-stimulation paradigms where electrodes are placed in various structures and animals are allowed to control the stimulation. In such studies most areas of the limbic system support self stimulation. Lateral hypothalamic nuclie is one structure often cited in these experiments. It should be recalled that the lateral hypothalamus is the feeding center. During food deprivationself stimulation here increases. Self stimulation also occurs with electrodes in the posterior hypothalamus. In addition the septal nuclie and the nucleus accumbens also support self stimulation. Just what do these areas have incommon? One answer may be the medial forebrain bundle which carrries noradrenergic fibers from the locus cereuleus to the anterior limbic system (septum and N. Accumbens). In addition this pathway carries some of the mesocorticolimbic dopmainergic fibers from the ventral tegmental area. Stimulation of the VTA results in self stimulation similar to hypothalamic and septal stimulation. Of interest are reports that dopaminergic neurotransmission underlies much of the self-stimulation. Drugs such as cocaine and amphetamine are believed to use this system to cause reinforced drug use or addiction. Reward and reinforcement acts to facilitate behavior patterns. This implies long term changes in neuronal responsivity. Obviously the processes are complex. In general long term changes are the result of induced changes in connectivity. One of the processes of such changes is known as kindling. The kindling paradigm basically occurs when subthrshold stimuli are repeated over time. The stimuli may be pharmacologic or electrical. The areas most sensitive to kindling also recieve dopaminergic innervation. In the kindling paradigm repeated subthreshold stimulation results in a gradual acquisition of afterdischarges and subsequently frank seizures. If kindling continues further the animal will develop autonomous seizures. It has become a useful model to develop anticonvulsants. Of interest is the fact that cocaine is one of the most potent pharmacologic kindling agents. The changes with kindling are long term suggesting that kindling or kindling like processes underlies the processes of memory and learning. An analogous finding is behavioral sensitization which occurs with
stimulant drugs. In this condition the pharmacologic response to stimulants increases.
"More for less". It has been suggested that such sensitization is similar to
kindling. Memory and learning is believed to be a part of this process. Users have
physiologic responses which accompany "craving" for stimulants. Such responses
may be initiated by cognitive processes which accompany the use of the drug. In other
words, the physiologic responses begin with considering the use of cocaine. This can be
demonstrated by giving addicts a heirarchy of sensory stimuli which accompany their drug
use. These studies will increase our knowledge of drug abuse, craving, and reward
motivated behaviors. In addition it will aid in our understanding disorders of reward
motivated behavior such as depression where well known changes in reward motivated
behaviors is seen. Serotonin; synthesis and release
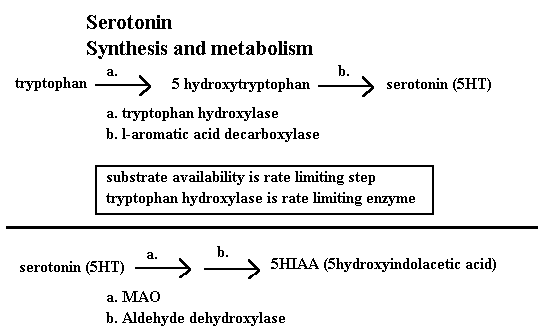
Serotonin is an indolamine monoamine neurotransmitter. The synthetic pathway is analogous to the catecholamines in many ways. An important distinction is that the rate limiting step is the uptake of tryptophan into the neuron. Tryptophan availability is the actual rate limiting factor in the intact animal. Tryptophan crosses the blood brain barrier via an active transport mechanism in competetion with other neutral amino acids such as leucine, lysine, and methionine. The activity of this transport mechanism is facilitated by the presence of insulin and glucose. Another interesting aspect of this system is the fact that tryptophan is one of the few amino acids which is bound in the plasma to any significant degree. The actual binding site is the fatty acid binding site of the albumen. This system allows a multitude of factors to ultimately influence the rate limiting step in serotonin synthesis. For example anything which increases free fatty acids would displace the tryptophan and thus increase the percent free which is able to cross the BBB. An example of such events include any acute stressor which increases glucocorticoid response, exercise, and acute alcohol consumption. The metabolism of serotonin is primarily done by MAO. The prinicple
metabolite is 5HIAA. The same statements concerning the CSF measurement of MHPG and HVA
applies to 5HIAA. Serotonin receptors There are three basic types of serotonin receptors; 5HT-1, 5HT-2, and
5HT-3. The 5HT-3 receptor is present in the area postrema which stimulates emesis. The
5HT-1 and 5HT-2 receptors are of a greater interest for psychiatry. The 5HT1 receptors
have been subtyped by DNA cloning and differential pharmacology into four major subtypes.
The most important is the 5HT-1a which is located in the Raphe and Hippocampus. This
receptor is implicated as an autoreceptor which modulates 5HT release from presynaptic
neurons. In addition the 5HT-1a receptors are G-protein linked and have been
implicated in thermoregulation, arteriolar vasomotor responses, hypotension, sexual
behavior, and possibly sleep. The 5HT-2 receptors are located throughout the cortex
and have been implicated in platelet aggregation, vasomotor contraction, head twitches,
and possibly sleep. Pharmacologically these receptors are important and are affected by a
wide variety of pharmacologic agents including butyrophenones, and phenothiazines. The projections of the serotonin system .
Note that they arise from the dorsal raphe and the raphe magnus. These nuclie are situated to observe ascending sensory input and act to facilitate information processing. One example of this is in the slow wave sleep. We will discuss this later when we discuss sleep and sleep disorders. Other implications of the sensory input into these nuclie are the concepts of sensory gating and directed attention. Recall that the Locus Ceruleus is important is arousal and vigilance. Vigilance is a state of increased arousal. Vigilance is necessary for focus or directed attention but not suffecient. Focused attention requires that incoming sensory information be given a priorty according to importance. The process of habituation occurs if stimuli is not reinforced. A lack of directed attention can appear as impaired concentration. Disrution of the normal serotonergic tone in animals can affect their
exploratory behavior. Animals seem to endow meaningless stimuli with relative behavioral
importance. That is they seem to have perseverative responses in serotonin
defiecient states and behavioral over activity in states of serotonin excess. It has been
suggested that some patients with neuroleptic resistant psychosis have a dysregulated
serotonin system. Some of the effects of atypical neuroleptics are thought to be mediated
through a 5HT antagonism. It would seem that these agents restore a proper sensory gating
and ability to direct attention Acetylcholine, synthesis and release Acetylcholine is one of the oldest and best understood neurotransmitters. This is likely because of its actions at the neuromuscular junction and the accessibility of this site to study. Unfortunately the ability to study the central cholinergic system is relatviely new. Unlike the catecholamines acetylcholine is a relatively simple structure with no aromatic rings. The ability to measure acetylcholine in body fluids had to await the development of better assays in the late 60's and 70's.
The synthesis of acetylcholine is relatively simple and straightforward. It is sythesized from acetylCoA + choline by an enzyme known as choline acetyltransferase (CAT). This enzyme is contained only in cells which synthesize acetylcholine. It is a marker enzyme which identifies cholinergic cells. Recall that Acetyl CoA is derived during the metabolism of glucose. Choline is derived from deitary sources and from phosphatidylcholines. After acetylcholine is released into the synapse it ultimately undergoes hydrolysis to release the choline and acetate. The choline is taken back up into the presynaptic neuron about 1/3 to 1/2 of the time. The choline can then be used ot resynthesize acetylcholine or can be used to synthesize phospholipids which can be used as stores of choline. The diagram below describes the points of pharmacologic interventions. Acetylcholine is metabolized by cholinesterases. There are several types the two most often referred to in reference to acetylcholine metabolism are acetylcholinesterase and butyrylcholinesterase or pseudocholinesterase. In the synapse the cholinesterase seems to be intimately related to the actual receptor. This enzyme is inhibited by physostigmine and organophasphates. Acetylcholine receptors There are two major types of cholinergic receptors based on differential binding. The first type is the nicotinic receptor and the second is the muscarinic receptor. DNA cloning has identified five subtypes of muscarinic receptors. All of these muscarinic receptors are G-protein linked. The muscarinic subtypes also seem to have a differential distribution in mammalian brain. The nicotinic receptor is a ligand gated channel composed of five subunits. It is primarily the responsible for the peripheral effects of acetylcholine at the autonomic ganglia and the neuromuscular junction. There may be some nicotinic receptors centrally but their contribution is uncertain. The nicotinic receptor is composed of 5 subunits. Of interest is the finding that injection of one of the more lipophilic subunits into the rabbit results in a syndrome indistinguishable from myasthenia gravis. This has confirmed suspicions that MG is an autoimmune disease. Nuclear medicine techniques are able to preferentially bind the
muscarinic receptors with an agent known as dexitimide and QNB. Studies are underway for
evaluating the sensitivity of these agents in the evaluation of Alzheimer's disease. Acetylcholine projections
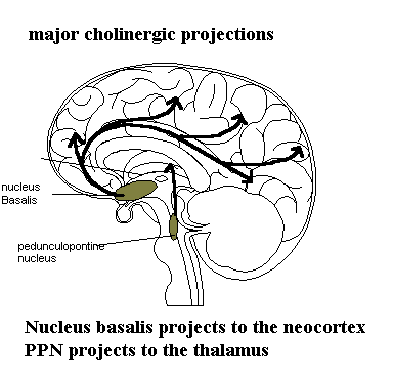
The chart below illustrates the major central projections of the cholinergic system. In essence there are three important cholinergic systems for neuropsychiatry. The first is the nucleus basalis of Meynert. This nuclie sends projects to the cortex. It is one of the nuclie which is impaired in senile dementia of the Alzheimer's type. It is suggested that this nuclie plays a role in learning and memory. In addition the cholinergic neurons in the basal forebrain are involved in cognitive intergration of vegative and motivationally relevant information. We will discuss this in more detail when we discuss the dementias. The second system arises from the brainstem. This system sends cholinergic fibers to the midbrain and thalamus. It is suggested that this system is related to sleep-wake rhythms and turning on REM. It is felt that this group of neurons acts as a sensory filter in some way. It has been shown, (here at UAMS) that schizophrenics has a less CAT in the pontine cholinergic nuclie. The meaning of this is uncertain. It is of interest that anticholinergic agents are well known psychotomimetics in high doses. The third group is the cholinergic neurons in the basal ganglia. These
will be discussed in more detail in the lecture on movement disorders. Gamma amino butyric acid, synthesis, storage and release GABA was identified in the mammalian brain in 1950's. It is believed to be the major inhibitory neurotransmitter in the brain. It is this role which is of interest to the neuropsychiatrist. GABA is synthesized from Glutamate as shown below. The marker enzyme is Glutamic acid decarboxylase (GAD). GAD is a pyridoxal cofactor dependent enzyme. A congential form of B-6 vitamin deficiency is known to predispose to seizures which are B-6 responsive. Glutamate is a pivotal amino acid in the brain. It is dervied from alpha keto glutarate which is one of the intermediates in the Krebs cycle by way of the addition of an amine group. Glutamate also undergoes transamination to form glutamine by addition of another amine group. Glutamine then proceeds to the liver where it is deaminated to regenerated glutamate which then returns to the brain. This is brain's nitorgen cycle. In situations where the liver is unable to deaminate the glutamine the brain must obtain glutamate by draining the Kreb's cycle intermediates. This in turn begins to impair cerebral energy metabolism. Following release GABA can be taken back up by the neurons or by
astrocytes. It appears that the release of GABA is also under autoreceptor control. GABA
is metabolized by the enzym GABA transaminase (GABA-T) to form succinic acid semialdehyde.
Succinic acid semialdehyde is metabolized further to form succinic acid which is also a
Kreb's cycle intermediate. GABA-T is inhibited by valproic acid. This is the basis for the
belief that valproic acid is GABAergic. There are other alternative pathways for GABA
metabolism. GABA receptors There are two basic subtypes, GABA-a and GABA-b. GABA-a is the most prevalent in the mammalian brain. The GABA-a receptor is similar to acetylcholine receptor in that it is related to an ion channel. In the case of GABA-a it is the chloride ionophore. Binding of GABA to this receptor increases the permeability to chloride ion which causes a hyperpolarization of the neuron or inhibition. The GABA-a receptor has four basic subunits, 2-alpha and 2 beta peptides which surround a chloride channel. There are three basic binding sites on this complex. The first is the GABA site. The second is a benzodiazepine site. The third is in the channel and is essentially a barbiturate site. Binding to the benzodiazepine site can have three effects, agonism, inverse agonism, or antagonism. The typical anxiolytic and sedative hypnotic agents such as diazepam and lorazepam act as agonist at these receptors. Their binding increases the affinity of the GABA binding site for GABA. This results in an increase in Cl- influx. Inverse agonism occurs with the beta carbolines. These agents act in the opposite direction as the agonist. Binding of these agents reduces the influx of Cl- below the baseline state. Clinically this is accompanied by anxiety. The antagonist such as flumazenil act to displace the agonist and inverse agonist without a direct effect on the chloride channel it's self. There is much speculation and a little evidence for an endogenous benzodiazepine ligand. It has been found to be increased in some metabolic conditions such as hepatic encephalopathy. In fact there are case reports of the use of benzodiazepine antagonist in the treatment of hepatic encephalopathy. Some patients have been noted to become less somnolent and more oriented. It remains to be seen whether flumazenil will have any use beyond reversing benzodiazepine overdoses.Clinically agents which are GABA-A agonist are generally anticonvulsant in activity as well as muscle relaxants. The GABA-b receptor is a G-protein related receptor which is distinct from the GABA-a sites. The highest concentrations of GABA-b receptors is in the interpeduncular nuclie and cerebellum. It appears that one of its prinicple effects is to increase the efflux of K+ from the cell. This would result in a hyperpolarization. Pharmacologically baclofen is considered a GABA-b agonist. The principle effect of GABA-b agonism is muscle relaxation. A significant relationship of dopamine and GABA exists. In general GABA
acts to reduce the firing of the dopaminergic neurons in the tegmentum and substantia
nigra. It forms the basis for the use of benzodiazepines as augmentation strategies in the
treatment of psychosis. In addition benzodiazepines may be helpful in cases where there is
an over activity of dopamine in the motor striatum such as Huntington's Chorea or Tardive
Dyskinesia. It is believed that they act by increasing the feedback inhibition. The
feedback inhibition from the GABA neurons of the globus pallidus and putamen to the
dopaminergic neurons of the substantia nigra is an important modulating force on the
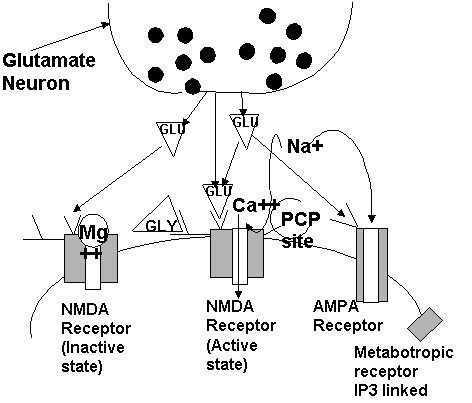
activity of the dopamine neurons. Excitatory amino acids Glutamate is considered the principle excitatory amino acid in the CNS. Its role in cellular metabolism is well known. Glutamate is distributed widely throughout the neuroaxis. Regions in which it seem particularly important include the granular cells of the cerebellum, the pyramidal cells of the hippocampus, the Betz cells of the motor strip, and the projections of the frontal lobe to the basal ganglia. One of the major difficulties in recognizing the role of glutamate as a neurotransmitter was the fact that there is only a small percentage of the glutamate present in synaptic vesicles. The vast majority of glutamate is present as part of intermediary metabolism. The ability to define the role of glutamte came as a result of finding receptors for glutamte and labeling them with various analogs. There are four glutamate receptor subtypes of importance. More will undoubtably be found and the nomenclature may change in the next few years. The four receptor subtypes include the NMDA receptor, the quisqualte receptor (also called the AMPA receptor), the kainate receptor, and the metabotropic receptor. A common feature of these receptors is depolarization of the membrane potential. In addition there are some special characteristic which make this system interesting. First the receptor systems seem to have a degree of cooperativity. By this I mean that they function most completely as a group. The diagram below illustrates these resceptor systems. Note that the NMDA receptor is present in an inactive state with a Mg++ ion blocking the calcium channel. For the NMDA receptor to become active the Mg++ must leave the channel site. This is accomplished by membrane depolarization brought about by the other glutamate receptors. Once this is done and the NMDA receptors are active a process known as excitotoxicity occurs. This may provide the basis for kindling and other longterm changes which result in neuronal specialization. Excitotoxicity is due to the opening of the calcium channels which results in an increase in free calcium. The degree of damage seems to be limited in most situations by an energy dependent mechanism of binding the calcium. In situations of ischemia or hypoglycemia there is a loss of this system and an excessive excitotoxicity. Neuroscience has attempted to finding specific NMDA receptor blockers to protect against ischemic damage. Some of the compounds found to date include the benzomorphans, MK801, and phencyclidine. Each of these compounds has psychotomimetic activity. This has limited there utility to date. Focus is no on blockers at other domains of the molecule such as the calcium channel itself. Another potential site suggested by the diagram is the glycine site. Glycine acts as an allosteric facillitator. Binding of glycine increases the ability of glutamate to activate the channel apparently. It is unclear if this is the site that Nitric oxide acts.
|


.GIF)
.GIF)
.GIF)
.GIF)